

0 引言
随着对生态环境的日益重视,有关城镇化所带来的环境问题及其对生物多样性影响的研究迅速增加,研究对象涉及到昆虫、植物、鸟类、哺乳类[3,4,5,6]等。相关研究可分为2类: 一是从空间或时间方面着手,以其为变量,设置城镇化的不同梯度,并对不同梯度内的生物种群变化进行记录,分析城镇化对其造成的影响[7,8,9,10]; 二是采用遥感解译与实地调查相结合,或是利用物种分布模型来模拟物种的潜在生境,分析栖息地的动态变化,结合同时期生物种群变化,分析影响生物多样性的原因[11,12,13,14]。2类研究,前者将城镇化作为一个整体变量,研究其对生物多样性的影响,略过栖息地变化这一过程; 后者则是注重于栖息地变化的影响,忽略了城镇化这一起因。城镇化是引起土地利用结构变化的重要原因,而物种栖息地被占用及其景观结构的变化又是生物多样性丧失的最直接的威胁之一[15],因此,城镇化进程对生物多样性的影响是大众所认同的。然而,现有研究中少有将城镇化与物种栖息地变化动态过程相结合,对两者间的相关性进行具体的分析。
1 研究区概况及数据源
1.1 研究区概况
洞庭湖地处湖南省北部、长江中游南岸(28°30'~30°20'N,110°40'~113°10'E),是我国第二大淡水湖和长江流域重要的调蓄湖泊。本文选取洞庭湖流域湖南省境内的岳阳、益阳、常德3市为研究区,包括共26个县级行政单元,总面积约4.54×104 km2,如图1所示。湖区周边地形为一碟形盆地,以洞庭湖为核心,向东、南、西三面过渡为河湖冲积平原、环湖丘陵岗地、低山。湖区归属亚热带季风气候区,温暖湿润、雨热充足,年平均温度为17 ℃,年平均降水量为1 200~1 450 mm。该区所特有的生态环境孕育了丰富的自然资源,是多种珍稀动物的栖息地。
图1
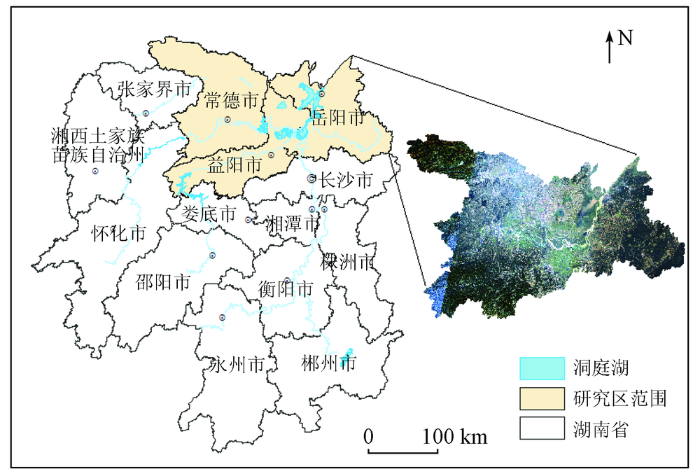
1.2 数据源及其预处理
研究所使用到的数据源主要有遥感影像数据、数字高程模型(digital elevation model,DEM)和矢量地图数据。其中影像数据为美国陆地资源卫星系列(表1),共5期35景,考虑到湖区水位的季节性变化,所获取数据的月份范围为该年的10—12月间; 其与DEM来源于地理空间数据云和美国地质勘探局官网,矢量地图数据来源于国家基础地理信息中心。后续过程中所涉及到的归一化差值植被指数(normalized difference vegetation index,NDVI)、比值植被指数(ratio vegetation index,RVI)、坡度等数据通过波段运算、空间分析等方式得出。
表1 遥感影像数据信息
Tab.1
| 年份 | 传感器 | 行列号范围 | 获取时间 |
|---|---|---|---|
| 1995年 | Landsat5 TM | 123~125,39~41 | 9月23日,12月5日,11月17日,12月28日 |
| 2000年 | Landsat5 TM | 123~125,39~41 | 10月06日,11月07日,12月25日 |
| 2005年 | Landsat5 TM | 123~125,39~41 | 9月09日,11月18日,12月13日 |
| 2010年 | Landsat5 TM | 123~125,39~41 | 11月12日,12月05日,12月21日,11月10日 |
| 2015年 | Landsat8 OLI_TIRS | 123~125,39~41 | 10月25日,10月16日,12月26日 |
2 研究方法
2.1 基于CART获取规则的决策树分类
分类与回归树(classification and regression tree,CART)是基于遥感影像数据及其他空间数据,通过专家经验总结、数学统计和归纳方法等来获得分类规则并进行遥感分类的方法,其基本原理是通过对训练数据集进行循环分析来形成二叉决策树结构[19]。根据研究区具体情况及研究内容,参考土地利用现状分类表,建立了研究区的分类体系,将研究区用地分成了林地、农田、建设用地、水系、滩地5类,其中滩地含泥滩地、芦苇滩地和苔草滩地3个亚类。参考地物类别的反射率特征选取影像Band1—Band7(2015年Band1—Band8),ISODATA分类结果,NDVI,RVI,DEM,坡度这12个分类特征参数(2015年为13个)进行波段合成,结果作为CART决策树分类对象,该波段组合可较好地对山体阴影、水体及3类滩地进行区分。
在ENVI中运行CART算法,获取决策树文件,并对输出结果进行聚类、筛除、主/次要分析等分类后处理,得出最终的洞庭湖区LUCC分类结果。
采用参考高分辨率的Google影像,随机选取各类样本点来与分类结果进行混淆矩阵计算的方法来进行分类精度评价,其中的主要评价参数有总体分类精度和Kappa系数。
2.2 扩张强度分析
将研究单元细化为网格大小,可更细致地反映研究区域的扩张差异,本文中选用城镇扩张强度指数(urbanization intensive index,UII),以网格为单位来表征区域内建设用地扩张的剧烈程度,其计算公式为:
式中:
2.3 鸟类生境适宜性分析
MaxEnt模型是一种物种分布预测模型。它将物种与其生长环境视为一个系统,通过计算系统具有最大熵时的状态参数确定物种和环境之间的稳定关系,并以此估计物种的分布[20,21]。它在形式上与Logistic回归模型相似,根据最大熵原则获取参数值估计,主要运作原理为利用已知物种分布点和相关环境变量,根据算法推算出物种的生态需求,将运算结果投射到不同的空间和时间来预测物种的潜在分布[22,23]。本文中参考洞庭湖自然保护区的鸟类监测报告及国家红色保护鸟类名录[24],选取了洞庭湖区白鹤、东方白鹳、中华秋沙鸭等国家Ⅰ、Ⅱ级保护鸟类共14种,地形、植被、气候、人类干扰等环境因子35种[24],通过MaxEnt模型来对洞庭湖区各鸟类生境分布及适宜性进行了分析。
3 结果与讨论
3.1 分类精度评价
精度评价结果如表2所示。研究区5期影像数据解译的总体精度都在94%以上,Kappa系数均大于0.92,其中分类精度最高为2015年,其总体精度为97.29%,Kappa系数为0.96。由此可见,整体分类结果较为精准,与普通的监督/非监督分类方法相比较,总体精度在整体上提升了8~13百分点,Kappa系数提升了0.09~0.15,在山体阴影及3类滩地的细分识别方面有明显提高。
表2 影像分类精度统计
Tab.2
| 年份 | 总体精度/% | Kappa系数 |
|---|---|---|
| 1995年 | 96.46 | 0.95 |
| 2000年 | 94.01 | 0.92 |
| 2005年 | 96.73 | 0.94 |
| 2010年 | 96.01 | 0.94 |
| 2015年 | 97.29 | 0.96 |
3.2 城镇扩张时空分异特征
3.2.1 城镇扩张时间异质性
城镇扩张的最直观表现为城镇建设用地规模在空间上的不断扩大,由建设用地面积统计(图2),可知在20 a间,洞庭湖区建设用地呈持续增长状态; 在不同历史时间段,扩张趋势有所不同,其扩张速率呈先增加后减少趋势。建设用地总面积在1995—2015年间增加了808.61 km2; 在2000—2005年呈现明显增长趋势,由312.02 km2增长为649.89 km2,平均增速为65.77 km2·a-1; 2005年后,扩张速度逐步变缓。
图2
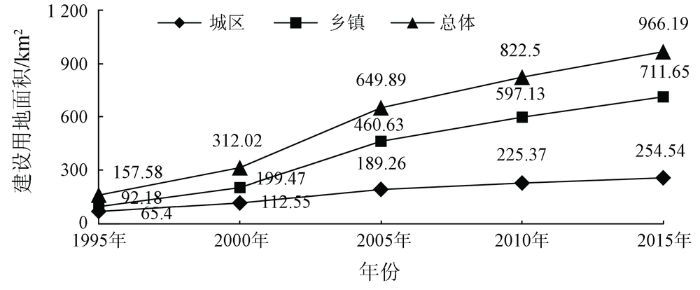
图2
1995—2015年洞庭湖区城镇用地面积统计
Fig.2
Statistics on construction land in Dongting Lake area from 1995 to 2015
将建设用地分为城区和乡镇2类分别统计扩张速度(表3),可发现,城区与乡镇建设用地的扩张总体趋势表现一致,皆由1995年的缓慢扩张开始,2000年后,进入迅速扩张阶段,并在2005年后,速度逐步减缓。而从面积增加量来看,存在较大差异,其原因是,城区与乡镇扩张速度差异在2000年后逐渐变大; 20 a间,城区建设用地的扩张面积为189.14 km2,而乡镇建设用地在此时间段内增长了619.47 km2,扩张面积为城区的3.27倍,因此,乡镇区域在这一时期经历了更为迅速的建设用地扩张过程。城区建设用地面积与统计年鉴中建成区面积相比较,其误差为2.92%,说明数据有较高的准确性。
表3 1995—2015年洞庭湖区城镇用地扩张速度统计
Tab.3
| 年份 | 城区 | 乡镇 | 总体 |
|---|---|---|---|
| 1995—2000年 | 9.43 | 21.46 | 30.89 |
| 2000—2005年 | 15.34 | 52.23 | 67.57 |
| 2005—2010年 | 7.22 | 27.3 | 34.52 |
| 2010—2015年 | 5.83 | 22.9 | 28.74 |
3.2.2 城镇扩张空间异质性
根据UII值将网格划分为逆扩张、无扩张以及低、中、高强度扩张5个层级,得到1995—2015年间洞庭湖区城区建设用地扩张强度分布图(图3)。经分析,在1995—2015年间网格总面积持续增加,低强度扩张为扩张主要类型,但不同时期城区扩张强度空间分布具有明显的差异性。在1995—2000年间,中高强度扩张网格较少,且多集中于市级的主城区小范围内,如岳阳市岳阳楼区、常德市武陵区、益阳市赫山区,距市级城区越远,扩张强度层级越低; 而在2000—2005年期间,中、高强度扩张网格数量增加,高强度扩张区域开始由市级中心向县级中心扩展,如临湘市、桃源县、津市市等离市中心较近的县级市中心也分布有高强度扩张网格; 至2005—2010年期间,中、高强度扩张网格进一步围绕市级城区中心扩大空间范围,并辐射到了石门县、安乡县、湘阴县等的边界县市; 经历过快速扩张的2010年后,扩张强度有所减缓,整体上高、中强度扩张网格大面积减少,除岳阳市的岳阳楼区、常德市武陵区以及澧县、湘阴县、汨罗市几个边界县级市以外,大部分空间都被无扩张网格覆盖。
图3
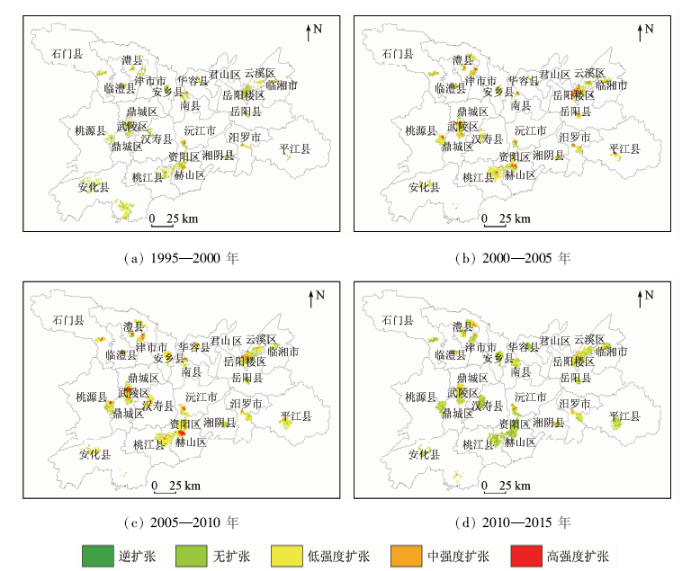
图3
1995—2015年间洞庭湖区城区建设用地扩张强度
Fig.3
Expand intensity of urban in Dongting Lake area from 1995 to 2015
以乡镇为研究单元进行扩张强度计算及统计分析的结果(图4)表明,20 a间,乡镇建设用地的扩张可以2010年为界分为2个阶段,在前一阶段,中、高强度扩张乡镇持续增加,为扩张加速期; 在后一阶段中乡镇扩张速度放缓。1995—2000年间,全域61.6%的乡镇进行着低强度扩张,而中、高强度扩张乡镇仅有10个,其中高强度扩张乡镇2个,为华容县城关镇和益阳市资阳区大码头街道。2000—2005年期间,研究区扩张速度提升,高强度扩张乡镇增加到7个,分别为云溪区的长岭街,汨罗市城关镇、新市镇,华容县城关镇、治河渡镇,赫山区朝阳街道,鼎城区武陵镇; 中强度扩张乡镇增加为28个,空间位置上多分布于高强度扩张乡镇周边; 但研究区的绝大部分乡镇保持低强度扩张状态,且3市的周边乡镇,多为无扩张状态。2005—2010年间,研究区内乡镇的扩张剧烈程度较之前5 a有所下降,扩张局势分布也有所不同; 前期无扩张的大部分边界乡镇在此时期内进入了低强度扩张状态,中强度扩张乡镇整体聚集于常德市安乡县内; 高强度扩张乡镇皆由其他层级转化而来,具体有君山区柳林洲街道,汨罗市汨罗镇,赫山区会龙山街道,沅江市万子湖乡和武陵区东江乡。2010—2015年间,扩张乡镇面积减少,但扩张剧烈程度与前一时期相比较有所回升,中强度扩张乡镇数增加为26个,空间上分布于常德市东北部和岳阳市南部,多由无扩张、低强度扩张类型转变而来; 高强度扩张乡镇有3个,分别为汨罗市新市镇,华容县城关镇,鼎城区武陵镇。20 a间,中高级扩张强度乡镇多出现于靠近国道、省道、高速、铁路等的区域,可推测交通为带动以上区域发展的重要因素。
图4
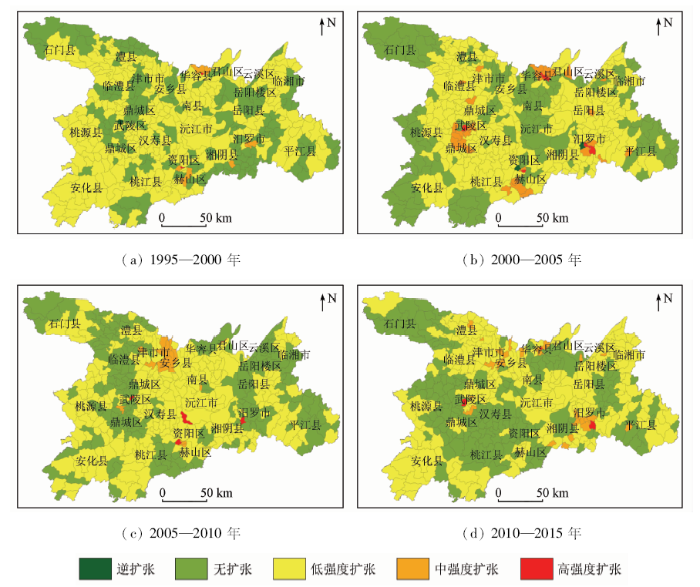
图4
1995—2015年洞庭湖区乡镇建设用地扩张强度
Fig.4
Expand intensity of townships in Dongting Lake area from 1995 to 2015
3.3 鸟类适宜生境时空分异特征
3.3.1 鸟类适宜生境时间异质性
根据鸟类的保护等级、濒危程度利用层次分析法来确定权重(表4),对MaxEnt运行得到的各鸟类生境适宜性值进行空间叠加,并依据值大小采用自然断点法划分为不适宜、较不适宜、适宜、最适宜4个层级,得出湖区的整体鸟类适宜生境分布,如图5所示。高适宜性程度区域与实际鸟类分布点情况相对比,确为鸟类聚集区域。对历年各适宜性面积进行统计(表5),可知1995—2015年间,鸟类生境的适宜和最适宜区域面积分别减少了144.07 km2和51.12 km2,而较不适宜区域面积在此20 a间增加了161.85 km2,研究区内鸟类生境在整体上呈现高等级向低等级、适宜区域向不适宜区域转化的趋势。各层级面积在不同时间段的变化情况也有所不同,如不适宜区面积在1995—2005年间逐年递减,2005年后面积又大量增加,适宜区变化情况与其正相反; 较不适宜区域在1995—2000年间面积大量增加后保持缓慢减少趋势,而最适宜区域则是从1995年开始面积持续减少,在2005—2010年间短暂回升后,再次回到逐步减少状态。
表4 鸟类权重赋值
Tab.4
| 保护等级 | 濒危程度 | 物种名 | 权重 |
|---|---|---|---|
| 国家Ⅰ级 | 极危 | 白鹤(Grus leucogeranus) | 0.305 7 |
| 濒危 | 东方白鹤(Ciconia boyciana)、中华秋沙鸭(Mergus squamatus) | 0.164 1 | |
| 易危 | 白头鹤(Grus monacha) | 0.087 7 | |
| 低危 | 黑鹳(Ciconia nigra) | 0.028 4 | |
| 国家Ⅱ级 | 易危 | 白枕鹤(Grus vipio) | 0.071 4 |
| 近危 | 小天鹅(Cygnus columbianus)、鸳鸯(Aix galericulata) | 0.035 7 | |
| 低危 | 灰鹤(Grus grus)、白额雁(Anser albifrons)、白琵鹭(Platalea leucorodia)、雀鹰(Accipiter nisus)、日本松雀鹰(Accipiter gularis)、松雀鹰(Accipiter virgatus) | 0.017 9 |
图5
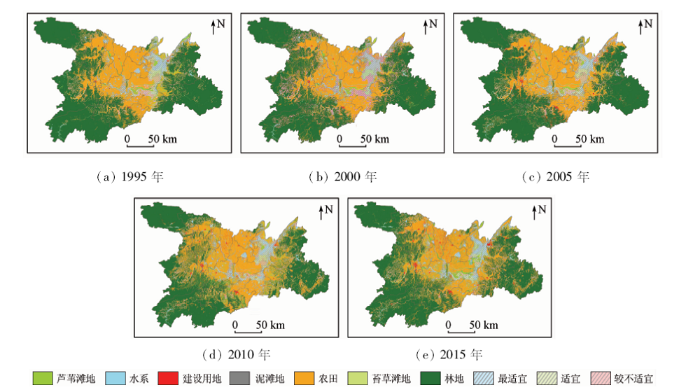
图5
1995—2015年洞庭湖区鸟类适宜生境空间分布
Fig.5
Spatial distribution of avian suitable habitat in Dongting Lake area from 1995 to 2015
表5 1995—2015年洞庭湖区鸟类适宜生境面积统计
Tab.5
| 年份 | 最适宜 | 适宜 | 较不适宜 | 不适宜 |
|---|---|---|---|---|
| 1995年 | 1 422.17 | 1 414.38 | 3 841.96 | 38 605.82 |
| 2000年 | 1 153.73 | 1 460.28 | 4 488.29 | 38 182.02 |
| 2005年 | 1 129.90 | 1 639.37 | 4 356.74 | 38 158.33 |
| 2010年 | 1 511.27 | 1 708.09 | 4 323.99 | 37 740.98 |
| 2015年 | 1 270.31 | 1 371.05 | 4 003.81 | 38 639.16 |
3.3.2 鸟类适宜生境空间异质性
根据研究区的网格单元,计算不同网格在各时间段的适宜性变化值,变化值为正则代表该区域向较高适宜性层级转变; 根据各个网格的变化值进行克里金插值,制作湖区鸟类生境适宜性变化空间分布图(图6)。由图6可知,在1995—2000年期间,南县、沅江市、岳阳县境内邻湖区域以及津市市与澧县交界处的区域(涔澹农场、如东乡)的适宜度都有较大程度的降低; 而适宜程度有所提高的则是离湖稍远的南县西部与安乡县邻界区域、岳阳县西部与华容县、沅江市邻界区域。2000—2005年间,适宜度继续降低的区域有东洞庭西部(岳阳县与华容县及其与沅江市相邻区域)、南县大通湖区域、桃源县西北部(漆河镇、黄石镇、龙潭镇)以及澧县西北部(盐井镇、火连坡镇、码头铺镇); 而适宜度发生较大提升的有湘阴县北部横岭湖区域、沅江市南洞庭芦苇场、南县大通湖以东以及平江县昌水沿河区域; 从整体空间范围上来看,东洞庭湖区适宜度持续降低,南洞庭湖区适宜度情况有所好转。2005—2010年间,整体上呈现出大范围水系周边适宜性程度所提升,而小型水系周围适宜度下降的景象。东洞庭、南洞庭沿湖区域以及平江县东部汨罗江以北区域的适宜度都有较大程度的提升; 而适宜度有所降低的区域为湘阴县横岭湖和岳阳楼区南湖以北区域、津市市北的涔澹农场和澧县北部牛奶湖东南岸区域以及桃源县泥窝塘乡北部的水库区域。2010—2015年间,除东洞庭西南岸的沅江市东部区域及西岸的华容县与君山区交界处的适宜度出现较大程度下降以外,其他区域整体情况都向适宜方向发展; 湘阴县北的横岭湖区域在这段时间内适宜程度持续提升,此外,适宜度有提升的区域还有东洞庭湖北岸(岳阳县新洲芦苇场、麻塘镇)以及临湘市东北部边界处、桃源县中部(泥窝塘乡北)的水库区域。
图6
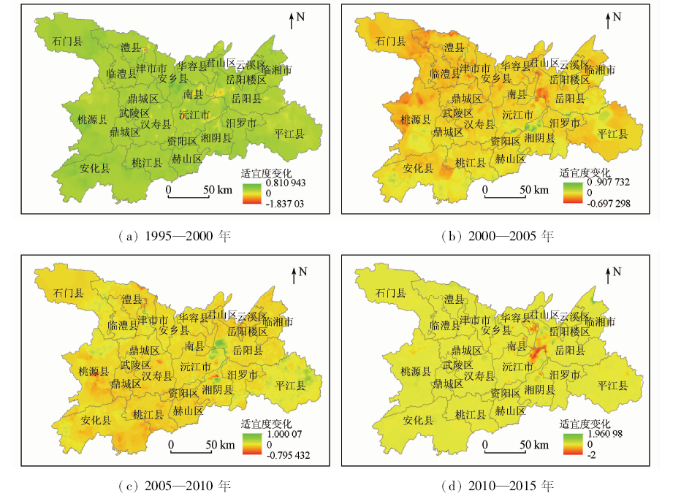
图6
1995—2015年洞庭湖区鸟类生境适宜性变化
Fig.6
Changes of avian habitat suitability in Dongting Lake area from 1995 to 2015
3.4 城镇化与鸟类生境变化相关性分析
计算研究区各时期城镇建设用地及栖息地各适宜程度面积的变化值,并对历年来的变化量进行多项式拟合,结果如图7所示。分析图中各类别曲线的变化趋势可发现,城区建设用地的扩张与各适宜程度面积的变化都没有出现明显的相关性,而乡镇建设用地面积的变化趋势与不适宜区域面积的变化趋势较为同步,且与最适宜生境面积的变化趋势呈相反趋势,说明乡镇建设用地面积的变化与不适宜区域的变化正相关,与最适宜区域的变化负相关,乡镇建设用地的扩张使得湖区鸟类栖息地的适宜度降低。从空间扩张方面解释,可能是由于城区的城镇扩张多为围绕中心城区向外蔓延,离鸟类栖息地的空间距离较远,因此并不能对栖息地环境造成直接的相关影响; 而乡镇距鸟类栖息地空间距离较近,且建设用地的扩张多较杂乱无序,以影响周边环境的水体、植被,改变周边环境景观格局为代价,而这些因素皆能直接影响鸟类栖息地适宜程度。
图7
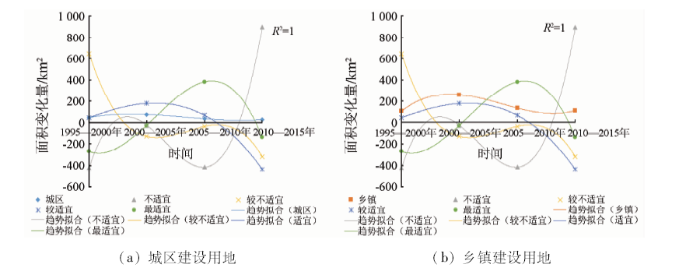
对各时期适宜程度发生明显转变的区域与乡镇建设用地扩张强度图进行比对可以看出,在空间范围上适宜度降低区域多为低强度扩张区域,适宜度提升区域为无扩张区域,而其他2类没有表现出与适宜性变化的空间相关。考虑到中、高强度扩张区域在空间位置上多靠近市区中心周围,此结果也可补充说明乡镇建设用地的扩张可影响鸟类栖息地适宜程度。在1995—2005年间,近湖区域的适宜度大面积降低,其内在原因大致为这一时期内湖区的大肆围垦、泥沙淤积、杨树种植和发展化工业,造成了水体面积的大量缩减以及湖区湿地环境的污染和破坏,而泥沙淤积问题起源于上游区域的植被破坏,上述问题都与城镇化因素主导下的人类经济活动相关联。在2005年以后,主体湖泊周边的生境适宜度的持续提升,得益于政府部门对湖区环境的整治工作,退田还湖,封山育林,对造成污染的企业及杨树林都进行了治理,且2005年后,城镇化扩张速度也有所降低,湖区生态环境逐步好转。因此,从面积变化量方面以及鸟类栖息地适宜程度变化的内在成因方面来看,洞庭湖区的城镇化扩张都影响了湖区鸟类栖息地适宜性。
4 结论
本文利用所获取的Landsat系列多时相遥感影像数据,通过构建洞庭湖区影像解译体系,并基于CART算法获取决策树规则的方法,对洞庭湖区土地利用类型进行解译,以其为基础,进行了有关洞庭湖流域范围内城镇化扩张和鸟类适宜生境时空分异特征及其间关联性的分析,具体结论如下:
1)文中所构建的洞庭湖区影像解译体系及分类方法,降低了湖区山体阴影以及“同物异谱”、“异物同谱”问题的影响,分类精度较普通监督/非监督分类方法有所提高。
2)湖区建设用地呈持续增长状态,在20 a间,城镇建设用地的扩张面积为808.61 km2,但城镇扩张速率在2005年后逐渐降低。期间的1995—2005年时间段为研究区城区的快速扩张期,城区中、高强度扩张面积持续增加,但位置多集中于城市主城区范围; 2005年以后,县级市区也逐渐加入到快速扩张的队列中; 但在2010年后,全域扩张强度减缓。在1995—2010年的15 a期间,乡镇范围的中、高强度扩张区域逐年增加,且此类乡镇多有高速、国道、省道等经过,可推测出交通是带动其发展的重要因素。
3)1995—2015年期间,鸟类适宜和最适宜生境面积共减少了195.19 km2,较适宜区域面积有所增加,表明研究区的整体生境适宜程度在下降。从空间范围来看,在早期,靠近主体湖区的区域适宜度都有不同程度降低,在2000年后,南洞庭湖区域的情况开始好转,东洞庭湖大面积也在2005年后开始向高适宜性层级转变,在2010年后,研究区域整体情况改善,验证了湖区治理工作的成效。
4)从面积变化量回归曲线来看,乡镇建设用地面积的变化与不适宜区域面积的变化成正相关关系,与最适宜区域的变化成负相关关系,可看出乡镇建设用地的扩张使得湖区鸟类栖息适宜程度降低,空间范围的比对结果也可进一步佐证此结论。因此,为保护湖区鸟类的适宜生境,应对进一步推进对湖区乡镇建设用地的扩张的整体空间规划和管控以及湖区生态用地的保护工作。
受数据以及知识经验的限制,研究还存在以下不足: 分类精度检验样本根据高分辨影像确定,与实地调查相比较,可能会存在类别偏差,因此可能会使分类结果精度值偏高; 相关性分析中没有对城镇化与鸟类栖息地间的关联性给出具体的量化关系; 湖区气候变化、围垦、杨树种植和化工业等活动导致的水体面积的缩减以及环境污染破坏皆是影响鸟类栖息地适宜性的重要内因,文中将后几项都较笼统地归咎于城镇化进程中的人为活动。在下一步的工作中应针对湖区不同空间范围适宜度变化的内在可能诱因进行更细致的分析。
参考文献
基于RS和GIS的城镇化扩展时空特征及驱动力分析
[D].
Analysis of spatial and temporal characteristics and driving force of urbanization based on RS and GIS
[D].
生物多样性丧失机制研究进展
[J].
Advancements of the researches on biodiversity loss mechanisms
[J].
Urbanization effects on carabid diversity in boreal forests
[J].DOI:10.14411/eje.2003.015 URL [本文引用: 1]
Urban-rural and temporal differences of woody plants and bird species in Harbin City,northeastern China
[J].
Great tits and the city:Distribution of genomic diversity and gene-environment associations along an urbanization gradient
[J].
Adaptability and vulnerability of high flying neotropical aerial insectivorous bats to urbanization
[J].
Lizard species distributions and habitat occupation along an urban gradient in Tucson,Arizona,USA
[J].
A cross-continental look at the patterns of avian species diversity and composition across an urbanisation gradient
[J].
The phylogenetic and functional diversity of regional breeding bird assemblages is reduced and constricted through urbanization
[J].DOI:10.1111/ddi.2018.24.issue-7 URL [本文引用: 1]
Diversity and assemblage filtering in ground-dwelling spiders (Araneae) along an urbanisation gradient in Denmark
[J].DOI:10.1007/s11252-018-0819-x URL [本文引用: 1]
The influence of urbanization on butterfly diversity in Hefei,Anhui Province
[J].To investigate the butterfly diversity in Hefei, we selected three regions along a gradient of urbanization, namely urban, suburban and rural regions from March to October, 2010. A total of 1484 butterfly individuals were collected, belonging to 7 families, 28 genera and 34 species, among which Pieris rapae, Pseudozizeeria maha and Polygonia caureum were dominant species. Diversity index of butterfly varied with the three different urbanization regions. The diversity indices of butterfly in the three different urbanization regions were in order of rural site > suburban site > urban site. Temporally, the Shannon index and richness index at rural and suburban sites were the highest in July, while they were the highest in August in urban site. The Simpson index at urban and suburban sites was the highest in August, while it was the highest in July at rural site. The evenness index in the three different urbanization areas was the highest in August. Therefore, it was suggested that different measures should be taken to conserve and enhance the biodiversity based on the urbanization level and habitat type under rapid urbanization. ]]>
人为干扰下西洞庭湖湿地景观格局变化及冬季水鸟的响应
[J].
DOI:10.3724/SP.J.1003.2013.11090
URL
[本文引用: 1]

Populus spp.)林面积增加了9倍, 芦苇(Phragmites australis)面积增加了30.6%, 水域、草滩、泥滩地等天然湿地面积分别减少了46.4%、49.8%和39.8%。湿地生境改变、破碎化严重, 景观破碎度指数(landscape fragmentation index, LFI)从1.239上升到2.897。3种典型生境中越冬水鸟的种群结构、数量及分布特征表明, 恢复湿地是当前部分越冬水鸟的主要栖息场所, 但水鸟群落结构简单, 多样性指数较低(H'=1.866); 自然湿地因严重破碎化, 面积变小, 水鸟数量较少, 但多样性指数较高(H'=2.118), 且是黑鹳(Ciconia nigra)、白鹤(Grus leucogeranus)等珍稀濒危物种的栖息地; 杨树林的种植改变了原有自然湿地景观, 仅发现水鸟1种2只, 说明其已不适宜水鸟栖息。本研究表明, 西洞庭湖湿地景观的改变、生境破碎化导致其生物多样性降低, 杨树种植对自然湿地的侵占是西洞庭湖湿地生态系统退化的主要原因, 在今后的湿地管理中应严格控制杨树林产业的发展, 并逐渐将人工杨树林恢复为自然湿地。]]>
Disturbance-driven changes to landscape patterns and responses of waterbirds at west Dongting Lake
[J].
DOI:10.3724/SP.J.1003.2013.11090
URL
[本文引用: 1]
H'=1.866). (2) Fragmented natural wetland provided habitat for diverse wintering birds, including rare and endangered black stork (Ciconia nigra) and Siberian crane (Grus leucogeranus), thus rising the Shannon-Wiener index (H'=2.118) for the bird community in this type of habitat, although waterbird population numbers remained low. (3) Poplar plantation was, in general, not suitable for waterbirds with only two individuals of one species observed in this habitat during the field survey period. In conclusion, landscape pattern change and habitat fragmentation reduced biodiversity at WDL with the planting of poplar trees being the key driver of wetland ecosystem degradation. For future management, forestation should be restricted while restoration of natural wetland should be a high priority.]]>
Ecological modelling of the distribution of the lanner falcon Falco feldeggii in Sicily at two spatial scales
[J].
基于MAXENT生态位模型对内蒙古根河地区驼鹿生境适宜性评价
[J].
Suitability evaluation of moose habitat in Genhe area of Inner Mongolia based on MAXENT model
[J].
Effects of habitat fragmentation on biodiversity
[J].DOI:10.1146/annurev.ecolsys.34.011802.132419 URL [本文引用: 1]
洞庭湖区近30年土地利用/覆盖变化对湿地的影响分析
[J].
Study on land use/cover change impact on wetlands of Dongting Lake area for nearly 30 years
[J].
洞庭湖湿地生态系统服务功能价值评估
[J].
The evaluation of the ecosystem service value in Dongting Lake wetland
[J].
洞庭湖区湿地生态系统健康综合评价
[J].湿地与海洋、森林并称为地球三大生态系统,湿地退化速度远大于其他生态系统,从而使得湿地健康评价与分析成为湿地领域研究的重点内容之一。本文以生态系统健康及压力-状态-响应模型为研究方法,综合利用遥感、野外实测和社会统计等多源数据,建立一套湿地生态系统健康评价指标体系,集成利用遥感和地理信息技术建立湿地生态系统健康评价模型,实施湿地压力、状态、响应、健康分析,揭示了洞庭湖湿地生态系统健康状况的空间分布规律。研究结果表明:(1) 洞庭湖区31%的湿地面临人类干扰压力较大,19%的湿地状态较好,82%的湿地生态系统发生很大变化。(2) 湿地的总面积在人类活动的干预下逐渐缩小,湿地综合健康较差、一般和较好的区域分别占总面积的6%、75%和19%,缺乏健康状况最好及最差的区域。(3) 湘阴县、沅江县及岳阳县的湿地生态系统健康状况较好,益阳县较差,安乡县等12个县市一般。
Assessment and analysis of wetland ecosystem health in Dongting Lake
[J].
基于影像多种特征的CART决策树分类方法及其应用
[J].以扬州市宝应县为研究区,采用主成分分析法对研究区影像进行数据压缩和单波段数据增强,利用灰度共生矩阵分析第一主成分的纹理信息.运用基于CART算法的决策树分类方法,选用影像的光谱特征值、删值以及纹理统计量值为测试变量,并通过计算确定决策树的节点规则,提取影像中主要地物信息.将分类结果与单纯依靠光谱特征的监督分类法结果相比较,表明基于影像多种特征的CART决策树分类方法分类精度较高,尤其较好地提取了围网养殖区和建设用地.
CART-based decision tree classifier using multi-feature of image and its application
[J].
物种分布模型的发展及评价方法
[J].
The development and evaluation of species distribution models
[J].
生态位模型的基本原理及其在生物多样性保护中的应用
[J].
DOI:10.3724/SP.J.1003.2013.09106
URL
[本文引用: 1]
生态位模型是利用物种已知的分布数据和相关环境变量, 根据一定的算法来推算物种的生态需求, 然后将运算结果投射至不同的空间和时间中来预测物种的实际分布和潜在分布。近年来, 该类模型被越来越多地应用在入侵生物学、保护生物学、全球气候变化对物种分布影响以及传染病空间传播的研究中。然而, 由于生态位模型的理论基础未被深入理解, 导致得出入侵物种生态位迁移等不符合实际的结论。作者从生态位与物种分布的关系、生态位模型构建的基本原理以及生态位模型和生态位的关系等方面探讨了生态位模型的理论基础。非生物的气候因素、物种间的相互作用和物种的迁移能力是影响物种分布的3个主要因素, 它们在不同的空间尺度下作用于物种的分布。生态位模型是利用物种分布点所关联的环境变量来模拟物种的分布, 这些分布点本身关联着该物种和其他物种间的相互作用, 因此生态位模型所模拟的是现实生态位(realized niche)或潜在生态位(potential niche), 而不是基础生态位(fundamental niche)。Grinnell生态位和Elton生态位均在生态位模型中得到反映, 这取决于环境变量类型的选择、所采用环境变量的分辨率以及物种自身的迁移能力。生态位模型在生物多样性保护中的应用主要包括物种的生态需求分析、未知物种或种群的探索和发现、自然保护区的选择和设计、物种入侵风险评价、气候变化对物种分布的影响、近缘物种生态位保守性及基于生态位分化的物种界定等方面。
Ecological niche modeling and its applications in biodiversity conservation
[J].
DOI:10.3724/SP.J.1003.2013.09106
URL
[本文引用: 1]
Based on the environmental variables that associated with species’ occurrence records, ecological niche modeling (ENM) seeks to characterize environmental conditions suitable for a particular species and then identify where suitable environmental habitats are distributed in the space. Recently, ENM has been used increasingly in biological invasion, conservation biology, biological responses to climate change, disease spatial transmission, and variety aspects of ecology and evolutionary biology research. However, the theoretical background of these applications is generally poorly understood, leading to artifactual conclusions in some studies (e.g. niche differentiation during species’ invasion). In this paper we discuss the relationship between niche and geographic distribution and introduce the theoretical basis of ENM, along with relationships between the niche and ENM. Abiotic/biotic, historical and dispersal factors are three key elements that determine species’ geographic distributions at different scales. By using environmental variables derived from distributional records, ENM is based on observations that already include effects of biotic interactions, therefore ENM is used to characterize somewhere between the realized niche and potential niche, not the fundamental niche. Grinnellian and Eltonian niches are both manifested in ENM calibration, depending on the types of variables used to fit model, the natural spatial scale at which they can be measured, and the dispersal of individuals throughout the environment. Applications of ENM in understanding ecological requirements of species, discovery of new species or populations, nature reserve design, predicting potential invasion, modeling biological responses to climate change, niche conservatism, and species delimitation are discussed in this paper.
基于MaxEnt模型西南地区高山植被对气候变化的响应评估
[J].
Assessment of alpine mean response to climate change in Southwest China based on MaxEnt model
[J].
1990—2013年来洞庭湖区鸟类生物多样性热点区时空动态及变动机理
[J].
Spatio-temporal dynamic and mechanism of avian biodiversity hotpots in Dongting Lake region during 1990—2013
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}